
Mother of Thousands, Mother of millions!
By: Lionel Perez Valenzuela
That rarest plant!
certainly Kalanchoe daigremontiana, give that impression when one looks at it. With all those little "seedlings" growing on the edge of the leaves. It looks very strange. Each time one of these small seedlings (propagules) from the edge of the leaves off, is likely to be achieved a mature plant. This form of reproduction property of Kalanchoe daigremontiana is asexual and highly efficient.
An exotic and toxic plant!
The Kalanchoe is native to Madagascar, is extremely durable and has spread to many regions of the world. Reproduces very quickly (as it says its common name "mother of thousands") and is toxic because it contains a cardiac glycoside, the Daigremontianina . is common in some regions of the world who suffer livestock property poisoning by eating this plant.
How do plants reproduce?
A very basic level we can say that plants can reproduce sexually or asexually. In sexual reproduction, they are very different haploid cells called gametes. fusion of male and female gametes leads to the development of an embryo (zygotic embryo) and then to the formation of the seed. But in
asexual reproduction, new plants arise through specialized vegetative organs, consisting of somatic cells, such as tubers, rhizomes, stolons, bulbs, etc. Although also can do so by other methods such as natural somatic embryogenesis (somatic embryo).
Therefore plants can develop two types of embryos, zygotic embryos (from sexual reproduction) and somatic embryos (by asexual reproduction). The plants that developed from zygotic embryos possess characteristics of both parents, however those that are formed from somatic embryos are genetically identical your parent (clones). Recall that in plants embryos are bipolar. A pole has the apical meristem that give rise to leaves and stems and the other pole has the root meristem that give rise wing roots.
In certain plants is very common the formation of somatic embryos, including seeds. For example, citrus fruits, where both types of embryos are formed almost simultaneously.
How they grow and develop plants?
Plants and animals grow and develop, but they do it differently. Formation in plants of organs (organogenesis) is performed post-embryonic development (embryogenesis). So we could say that plants have two programs, one organogenic and embryogenic another.
embryogenesis and organogenesis are separate processes in plants.
When a plant reproduces in a sexual way, occurs egg fertilization (through pollination), we obtain the zygote, an embryo develops, the seed (the process of seed formation it is called, embryogenesis), the accessory structures and the fruit.
embryos in plants must go through a process of desiccation (water loss) essential for cellular metabolism stop and the seed is in a dormant state until it is reactivated. Therefore seed with its embryo is capable of withstanding until the fruit ripens, falls, breaks down, the seed is free and eventually (if they found an appropriate way) when in contact with water reactive metabolism begins to germinate and grow the seedling from the embryo (this process will be called organogenesis).
Consider for a moment how important it is that the seed pass through a dormancy period of apparent inactivity, because it can store and transport the seeds for planting in the right time.
What are meristems?
All plants have the same basic pattern of body schema, consisting of repeating units of stem and leaf. As we said before these units of stem and leaf, stem from the apical meristem, while roots grow from the root tips.
The outbreak apical meristem or bud (in English, shoot apical meristem or SAM, in its abbreviation), contains in its center, a group of stem cells, totipotent undifferentiated, which is self-renewing. As stem cells divide, daughter cells are pushed to the edge of the meristem, where clusters of these cells, they differ in leaf founder cells.
We can therefore say that the meristem tissues are formed by a group of totipotent cells, undifferentiated , which is self-renewing and generating other cell types that in turn will form the various tissues and organs plants.
Both apical meristem as the root, are generated during embryogenesis (formation of the seed), but do not participate in embryo development. The meristems are activated once the seedlings have germinated and remained active throughout the life of the plant, being a reservoir of new cells for organogenesis (development of stem, leaves, flowers).
totipotency in plant cells.
somatic cells of plants , unlike cells somatic animals have a remarkable feature , can regenerate a complete organism. This potential for regeneration is called totipotency.
This ability of plants is widely used to multiply asexually, for example from somatic embryos. So from a single plant in a laboratory, can be grown hundreds of plants genetically identical to the parent plant and repeat this process again and again.
Control of embryogenesis and organogenesis in plants.
varis genes have been identified master regulators of organogenesis and plant embryogenesis. One of the most important is the gene STM (Shoot Meristemless), a gene similar to the homeotic genes Knotted1 (English Knotted1-like homeobox). These genes generally are called genes KNOX1. We will see in another series of articles that homeotic genes control development in plants and animals. In this case when the STM gene is mutated, the apical meristem (SAM remember that shorten), not formed and therefore the plants can not grow and remain in the seed stage. Then we can say that the STM gene is important for organogenesis. Furthermore, when this gene is establishing he sobreeexpresa plants show growth buds ectopic (misplaced) in the leaves.
Genes essential for the formation of both types of embryos (somatic and zygotic) are the LEC1 (Leafy Cotyledon 1) and Fus3. If these genes are mutated the embryos stop their development, are intolerant to desiccation and therefore unworkable. When these genes are overexpressed in transgenic plants ectopically form somatic embryos from vegetative cells.
Interestingly, ectopic overexpression of either the genes that control organogenesis (STM) and embryogenesis (LEC1, Fus3), forcing the vegetative cells of genetically modified plants to form structures similar to the seedlings of Kalanchoe daigremontiana. So the researchers decided to study the activity of STM genes, LEC1 and Fus3 directly on the seedlings, so check directly which are the genetic programs involved in its formation.
plants of the genus Kalanchoe, an excellent experimental model.
the genus Kalanchoe plants show different strategies evolutive
1 - Species that reproduce only sexually and are not seedlings. Therefore zygotic embryos can form and have the LEC1 gene active.
2 - Species like the Kalanchoe daigremontiana only reproduce asexually and form seedlings at the edge of the leaves, constitutively. All these species have the LEC1 gene mutated, and possibly inactive or altered function. These plants can not form embryos (hence the seeds are viable).
3 - Species that can reproduce sexually (form viable seeds) and semiconstitutiva produce seedlings as well as in situations of stress. LEC1 gene active. These species like the Kalanchoe gastonis-bonnieri, which has all the reproductive strategies represent the ancestral evolutionary line.
4 - species that reproduce sexually, but that under conditions of stress can produce seedlings. LEC1 gene active.
Evolution asexual reproduction in Kalanchoe.
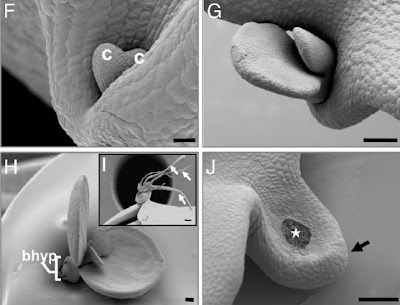
As we saw daigremontana Kalanchoe, is reproduced only asexually , seeds and embryos is not normal, but on the contrary, these seedlings develop very small (their chips seem cotyledons), which still attached to the mother plant, begin to form adventitious roots. At no time suffer desiccation, but they have their own transport system and even seem, are not embryos.
Why does this process of reproduction asexual? Well apparently some plants of this genus, have lost the ability to reproduce sexually, as they have the mutated gene LEC1 key in the formation of seeds and therefore have adapted to asexual reproduction. Or, conversely, may be that the mutation somehow LEC1 favors asexual reproduction.
most recent studies have shown that Kalanchoe daigremontiana plantlets form at the edge of the leaves, because both programs the organogenic and embryogenic work cooperatively on the sheet. These conclusions are based on three points:
1 - Kalanchoe daigremontiana STM gene can initiate a process of developmental change and generate a group of undifferentiated cells and competent as a meristem at the edge of the leaves (organogenic program).
2 - STM gene deletion prevents the development of the seedlings at the edge of the leaves.
3 - Both the LEC1 gene (mutated) as the Fus3 gene is expressed actively in the formation of the seedlings (embryogenic program), and therefore have been recruited into the organogenic program of development of seedlings, and would be responsible for some of the These unique features of somatic propagules (embryo-like structure).
The evidence provided suggests that asexual reproduction then it probably began as a process of organogenesis then recruited an embryogenesis program into the leaves, in response to the loss of the ability of sexual reproduction this genre.
important Why are these jobs?
The answer is simple, help us to understand how plants reproduce, evolve and adapt. Little-known phenomena and poorly understood at present.
addition there is enormous interest in the production of plants from somatic embryos, artificial seed technology, plant breeding endangered or displaced native. Propagation of fish with high commercial value, hybrid selection, reproduction studies of genetically engineered plants, sanitation, disease, etc, etc, etc. All these technologies could have a major impact on our ability to produce seeds and plants more resistant on the market value or food and therefore affect our ability to provide more and better food of plant origin is a growing population with an environment increasingly degraded.
Source: MP Helena Garces, Neelima R. Sinha (2007) Proc Natl Acad Sci USA, vol 104:15578-15583




